|
|
02 Features
Print To PDF
DisplayTitle Rediscovering Ancient Wheats
Authors Sabrina Geisslitz and Katharina A. Scherf1
Affiliations Department of Bioactive and Functional Food Chemistry, Institute of Applied Biosciences, Karlsruhe Institute of Technology (KIT), Karlsruhe, Germany
1 Corresponding author. Department of Bioactive and Functional Food Chemistry, Institute of Applied Biosciences, Karlsruhe Institute of Technology (KIT), Adenauerring 20a, 76131, Karlsruhe, Germany. LinkedIn: https://www.linkedin.com/in/katharina-scherf-96a44b83; KIT website: http://bioactivefc.iab.kit.edu
© 2020 Cereals & Grains Association
Abstract
CFWAbstract The ancient wheats spelt, emmer, khorasan wheat, and einkorn have been attracting more attention recently as consumer demand for products with health benefits and favorable nutritional compositions has grown. Furthermore, consumers are willing to spend more on regional, organic, and sustainably sourced and manufactured artisanal cereal products. Although the use of ancient wheats presents some challenges due to low yields, a tendency to lodge, and comparatively poor dough workability, it also presents multiple opportunities along the value chain to help safeguard and increase biodiversity and to create a variety of specialty products with excellent nutritional and sensory properties.
Trying to reach content?
View Full Article
if you don't have access, become a member
Page Content Wheat Cultivation in the 21st CenturyNutritional security worldwide is highly dependent on the production of cereals, which are grown on about 60% of agricultural land. In 2016, more than 2,800 × 106 tons of cereal grains were harvested worldwide, with maize (1,060 × 106 tons), wheat (749 × 106 tons), and paddy rice (741 × 106 tons) as the major crops (11). Wheat (Triticum spp.) includes the “ancient” (hulled) wheat species einkorn (T. monococcum L. subsp.
monococcum, diploid, genome AA), emmer (T. turgidum L. subsp. dicoccum, tetraploid, genome AABB), khorasan wheat (T. turgidum L. subsp.
turanicum, tetraploid, genome AABB, also known under the trademark Kamut®), and spelt (T. aestivum L. subsp. spelta, hexaploid, genome AABBDD), as well as the “modern” (free-threshing) wheat species durum wheat (T. turgidum L. subsp. durum, tetraploid, genome AABB) and common wheat (T. aestivum L. subsp.
aestivum, hexaploid, genome AABBDD) (Fig. 1). About 95% of the total wheat produced is common wheat, and about 5% is durum wheat. Spelt, emmer, and einkorn play only minor roles in terms of production and utilization.

The term “ancient wheat” is most often used to refer to spelt, emmer, khorasan wheat, and einkorn. In more general terms, “ancient grains” refers to populations of primitive grains that have not been subjected to any modern breeding or selection and, therefore, have retained characteristics of their wild ancestors, such as large individual variety, ear height, brittle rachis, and low harvest index (15). Ancient grains include small grains, such as teff, sorghum, and millet, as well as pseudocereals, such as amaranth, buckwheat, chia, and quinoa. In contrast, “modern wheats” are the result of ongoing breeding and selection with the aim of producing genetically homogeneous lines with stable and improved traits to ensure higher yields and productivity compared with “landraces” that have developed through natural and human selection but that are heterogeneous and well adapted to regional environmental conditions (33). Since the 13th century, einkorn and emmer have almost completely disappeared from agricultural fields. Einkorn is only cultivated in very small amounts in some regions of Austria, France, Germany, Hungary, and Italy. Although emmer only plays a minor role in Europe, it is more commonly cultivated in Ethiopia and India, especially in regions with harsh environmental conditions (18). Spelt was widely grown in southwestern Germany and other regions until the 19th century, when it disappeared almost completely because consumer preferences shifted toward white (refined flour) breads and away from whole grain breads and porridges. There are several reasons for the dominance of common wheat in modern agriculture. Ancient wheats are hulled, which means the grains are tightly enclosed by tough glumes (husks) that must be removed using a special dehulling treatment prior to milling. In addition, ancient wheats have long stalks and a tendency to lodge, even on nonfertilized soils, which can cause potential problems during harvesting. The yield for ancient wheats is also considerably lower (30–67%) compared with common wheat (19). In addition, doughs made with ancient wheat flours are typically difficult to handle, and breadmaking performance tends to be low (14). Ancient Wheats as the World’s First Domesticated Species
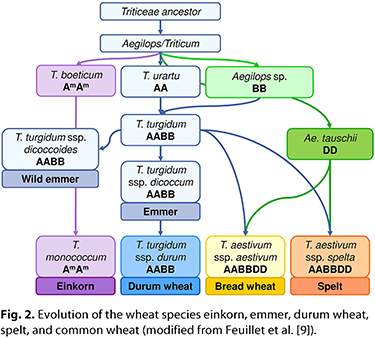 Wheat cultivation began about 10,000 years ago in the Fertile Crescent region formed by the Euphrates and Tigris rivers in what is now Turkey, Syria, and Iraq. During the Neolithic Revolution, humans evolved from societies of hunters and gatherers that moved from place to place to societies that built settlements and began farming in one place. The first wild ancestors of wheat were diploid wild einkorn (T. boeoticum, genome AmAm) and red wild einkorn (T. urartu, genome AA), both of which are separated by crossing barriers, as indicated by the superscript “m” used to differentiate both sets of the A genome (6). The name einkorn literally means “single grain” in German, because each spikelet contains only one grain. Although
T. urartu has never been domesticated,
T. boeoticum is the ancestor of domesticated einkorn (Fig. 2). Wild emmer (T. turgidum subsp.
dicoccoides, tetraploid, genome AABB) evolved from the spontaneous hybridization of
T. urartu and an unidentified
Aegilops species that contributed the B genome (21). Its domestication eventually led to the development of emmer, which is characterized by the presence of two grains per spikelet. Free-threshing emmer genotypes occurred spontaneously and subsequently were selected by humans for further breeding due to the absence of husks, eventually resulting in the creation of durum wheat. About 7,500 years ago, durum wheat was already being cultivated in the Fertile Crescent (22). Hexaploid wheats evolved from spontaneous hybridization of
T. turgidum species with goat grass (A. tauschii, diploid, genome DD). The origins of common wheat and spelt are not entirely clear, but it is most likely that durum wheat and
A. tauschii eventually evolved into common wheat, whereas spelt is a crossbreed of emmer and common wheat (3). Common wheat and spelt are readily interbred, and there are certain spelt cultivars that show a protein profile typical of spelt, while others display a profile that resembles common wheat (17). Wheat cultivation began about 10,000 years ago in the Fertile Crescent region formed by the Euphrates and Tigris rivers in what is now Turkey, Syria, and Iraq. During the Neolithic Revolution, humans evolved from societies of hunters and gatherers that moved from place to place to societies that built settlements and began farming in one place. The first wild ancestors of wheat were diploid wild einkorn (T. boeoticum, genome AmAm) and red wild einkorn (T. urartu, genome AA), both of which are separated by crossing barriers, as indicated by the superscript “m” used to differentiate both sets of the A genome (6). The name einkorn literally means “single grain” in German, because each spikelet contains only one grain. Although
T. urartu has never been domesticated,
T. boeoticum is the ancestor of domesticated einkorn (Fig. 2). Wild emmer (T. turgidum subsp.
dicoccoides, tetraploid, genome AABB) evolved from the spontaneous hybridization of
T. urartu and an unidentified
Aegilops species that contributed the B genome (21). Its domestication eventually led to the development of emmer, which is characterized by the presence of two grains per spikelet. Free-threshing emmer genotypes occurred spontaneously and subsequently were selected by humans for further breeding due to the absence of husks, eventually resulting in the creation of durum wheat. About 7,500 years ago, durum wheat was already being cultivated in the Fertile Crescent (22). Hexaploid wheats evolved from spontaneous hybridization of
T. turgidum species with goat grass (A. tauschii, diploid, genome DD). The origins of common wheat and spelt are not entirely clear, but it is most likely that durum wheat and
A. tauschii eventually evolved into common wheat, whereas spelt is a crossbreed of emmer and common wheat (3). Common wheat and spelt are readily interbred, and there are certain spelt cultivars that show a protein profile typical of spelt, while others display a profile that resembles common wheat (17). Crop domestication generally is characterized first by modification of key traits followed by subsequent selection, causing incremental changes in morphological and physiological traits (2). Wild wheats have small seeds with a hull and a rachis that shatters when the grain is ripe, so the seeds can disperse as widely as possible. The shattering rachis that is useful for plant reproduction presents a challenge for humans, however, because it is easier to harvest grains from the plant rather than from the surrounding earth. In addition to a rachis that does not shatter, spike size, growing season, plant height, seed dormancy, and grain size were useful traits for selection. Composition and Bioactive Components of Ancient and Modern WheatsThe composition of wheat grains typically consists of 56–74% available carbohydrates (mainly starch), 11–14% water, 8–12% protein, 2–13% dietary fiber, 2–7% lipids, and 1–3% minerals. Important minor constituents include vitamins such as the B vitamins niacin (15–55 mg/kg), riboflavin (1–2 mg/kg), thiamin (4–7 mg/kg), and pantothenic acid (7–17 mg/kg) (34). Wheat bread provides about 34% of the carbohydrates, 34% of the protein, 24% of the dietary fiber, and 13–32% of the B vitamins and minerals of the mean recommended daily nutritional intake in Germany (38) and many other countries with similar dietary habits. Bioactive components in wheat include dietary fiber; phytochemicals such as carotenoids and polyphenols, including phenolic acids, flavonoids and lignans; and vitamins. Flours made from smaller grains typically have higher contents of crude fiber, minerals, and other components that are enriched in the bran. The composition of grain constituents is not only affected by genetic background, but also by environmental factors, such as cultivation region, climatic conditions, and agronomic practices, in particular the type and amount of nitrogen fertilization. Increased nitrogen application leads to higher protein contents but also causes changes in protein composition, with higher proportions of storage proteins (gluten), especially gliadins, relative to nongluten proteins (27). Climatic conditions also impact grain composition, e.g., with positive correlations between phytochemical contents and the mean temperature during grain development and with negative correlations between water-soluble dietary fiber contents and temperature. Arabinoxylan, tocol, sterol, and alkylresorcinol contents are mainly determined by genetic factors, whereas phenolic acid, B vitamin, and betaine contents are influenced more strongly by environmental factors than by genetic background (32). The contents of major constituents, as well as minerals and vitamins, differ little between ancient wheats and modern common wheat and durum wheat (Table I).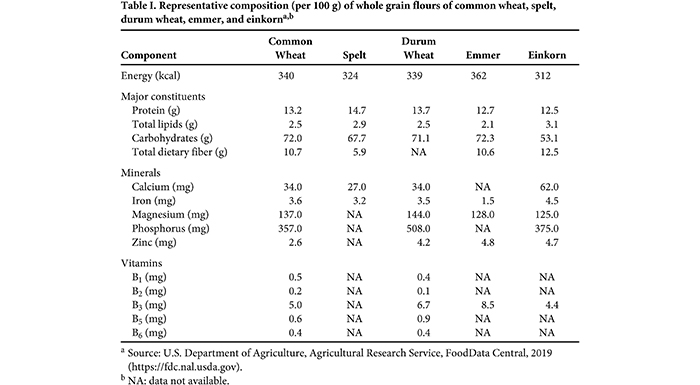
The same is true for most bioactive components, with the exception of the carotenoid lutein, which is higher in einkorn, emmer, and khorasan wheat compared with common wheat. However, durum wheat also has a high lutein content that has been increased by breeding, because the yellow color is desirable for pasta products (31). A recent study comparing the protein composition of 60 cultivars of ancient and modern wheats grown at 4 locations found that ancient wheats had higher protein and gluten contents than common wheat independent of location. With regard to protein composition, ancient wheats had lower glutenin contents compared with common wheat. Because high glutenin content is associated with good baking quality, this is most likely the main reason for the poor baking properties of many flours derived from ancient wheats (12). The Comeback of Ancient WheatsThere are several reasons why ancient wheats are currently experiencing a comeback. One of them is the global loss of biodiversity due to streamlined and uniform agriculture that relies on few species and, thus, leaves ecosystems vulnerable to pests and climate change. To counter this development, ancient wheats are important genetic resources for reintroducing genetic heterogeneity and providing special traits, such as disease tolerance, adaptability to a warming climate, and nitrogen use efficiency (23). For example, emmer and einkorn have higher nitrogen partial factor productivity and protein yield efficiency compared with common wheat, making them promising candidates for more sustainable agriculture with less nitrate pollution (12). Ancient wheats are especially well suited for organic agriculture with low agronomic input and often disadvantageous growing conditions. Another reason for the comeback of ancient wheats is that consumer trends are shifting away from industrially produced foods to sustainable, regional, and artisanal products that have been manufactured in ways that conserve resources. Ancient wheat products meet the increasing demand for individual consumption, differentiated offerings, and product variety. As a result, niche products are becoming more and more important in the marketplace. In particular, regional products are associated with attributes such as fresh, tasty, nutritionally valuable, healthy, and safe. Other aspects of these trends include the willingness of consumers to pay higher prices for regional products to support local food producers, contribute to fair pricing, reduce the ecological footprint, and promote biodiversity (1). In addition, ancient wheats have gained renewed interest because of the increasingly controversial debate in the media and social networks concerning wheat-related health risks. About 1% of the world population suffers from celiac disease, an immune-mediated inflammatory reaction of the upper small intestine that is caused by a combination of genetic risk, gluten consumption, and a certain unknown factor that initially triggers disease onset (20). Wheat allergies affect about 0.5% of the population (25), whereas the prevalence of non-celiac gluten sensitivity (NCGS) may be much higher at up to 6% (7). Celiac disease patients need to follow a strict gluten-free diet with a maximal intake of 20 mg of gluten/day. This means that these individuals need to eliminate wheat, rye, and barley products from their diet. In this context, it is important to emphasize that ancient wheats cannot be tolerated by celiac disease patients because they also contain gluten. In the case of wheat allergies, the treatment is strict adherence to a wheat-free diet, whereas it may be sufficient for NCGS patients to reduce consumption of wheat to 5–10% of their average daily intake (29). Recent surveys suggest that 20–30% of the Western population partially or completely removes gluten from their diets even though they do not have a medical diagnosis of any wheat-related disorder. Many of these individuals report that they tolerate traditional, artisanal baked goods made from ancient wheats better than common white wheat bread. It is currently unknown which specific components may be responsible for this difference in tolerability, but the underlying hypothesis is that ancient wheats and products made with them may contain fewer immunoreactive components. Potentially immunoreactive components in wheat, rye, and barley include gluten, alpha-amylase/trypsin-inhibitors (ATIs), wheat germ agglutinin, various other nongluten proteins, and fermentable, oligo-, di-, and monosaccharides and polyols (FODMAPs) (29). Gluten is known to cause celiac disease, but it is also a known allergen and may be involved in triggering NCGS. ATIs are plant-defense proteins that inhibit the digestive enzymes alpha-amylase and trypsin, thereby causing antinutritive effects. Already known as major wheat allergens (35), ATIs have been shown to activate the toll-like receptor 4 and trigger the innate immune response, as well as act as adjuvants of pre-existing inflammatory reactions (39). Although all ancient and modern wheats contain gluten, the load of known celiac disease-active amino acid sequences may vary among species. For example, due to the absence of the D genome, emmer, durum wheat, and einkorn do not contain the so-called 33-mer that is frequently described as the immunodominant peptide (28). Recent studies have shown that einkorn contains very low amounts of ATIs (13) and, therefore, may be a promising candidate for developing products with better tolerability for NCGS patients. Other in vitro studies have shown that einkorn has the lowest immunostimulatory activity, followed by durum wheat and emmer and then wheat and spelt (24,36,37). Taken together, there are more and more studies comparing bioactive components and immunostimulatory properties of ancient and modern wheats, but they often produce inconsistent results that range from clear differences between species to no differences. One important point for future studies seeking to clarify beneficial or adverse health effects is that the wheat samples used in testing need to be grown under the same conditions to minimize the confounding effects of environmental variability (30). Products Made with Ancient WheatsDue to their low yields, products made with ancient wheats will most likely never become suitable for the mass market, but they are well suited to rediscovery of traditional recipes and manufacturing techniques for creating specialty products. Ancient wheats are often used as whole grains in salads and soups, with emmer and einkorn used mainly for pasta and spelt used mainly for baked goods (5). Einkorn flour is usually difficult to utilize, because it forms a sticky dough with poor rheological properties upon addition of water. However, there are certain cultivars with high breadmaking quality that appear to be promising for further enhancement through breeding (4,12). Breads made from khorasan wheat had good sensory properties and loaf volumes that were almost as high as those made with common wheat (26). So far, the reintroduction of spelt products has been very successful, with an annual turnover of more than 1 billion euros across the product value chain in Europe (18). Spelt breads have a rich texture and flavor, and many traditional recipes, including long dough fermentation times, are being utilized to create an increasing variety of classical breads, fine pastries, and even pretzels. Although the functional and technological properties of gluten proteins in ancient wheats are typically inferior to those found in common and durum wheats, spelt also allows the production of pasta with satisfactory cooking quality. Pasta made with emmer even has improved sensory properties and a lower glycemic index compared with pasta made with durum wheat (8). Ancient wheat flours are also excellent for use in new specialty baked goods, crackers, snacks, flakes, muesli, and grits, as well as beer (10,16). ConclusionsThe rediscovery of ancient wheats offers new opportunities for farmers, millers, bakers, and consumers to create specialty products that are associated with good taste, enjoyment, health, naturalness, regional origin, and, often, organic agriculture. With their wide genetic diversity, ancient wheats are also valuable resources for crop biodiversity that may help improve nutrition security in light of climate change. Despite their low yields, ancient wheats are suitable for extensive agricultural systems in marginal regions under high-stress conditions where common wheat cultivars fail to thrive. Further multidisciplinary research efforts are needed to screen hundreds of ancient wheat cultivars; assess promising candidates for agronomic, processing, and nutritional properties; continue breeding efforts; and develop products with excellent nutrition and sensory qualities.
 Sabrina Geisslitz graduated with a degree in food chemistry from the Technical University of Munich (TUM) in 2015 and worked as a Ph.D. student at the Leibniz-Institute for Food Systems Biology at TUM. In 2019, she received her Ph.D. degree for her research on proteins in einkorn, emmer, and spelt, with a focus on technological properties and triggers of wheat-related hypersensitivities. The Cereals & Grains Association awarded her work with the Walter Bushuk Graduate Research Award for excellence in cereal protein chemistry. Currently, she works as a postdoctoral researcher in the group of Prof. Katharina Scherf at the Karlsruhe Institute of Technology (KIT). Her research studies the relationship between the structure and functionality of proteins in ancient and modern wheats using modern proteomics methods.
Sabrina Geisslitz graduated with a degree in food chemistry from the Technical University of Munich (TUM) in 2015 and worked as a Ph.D. student at the Leibniz-Institute for Food Systems Biology at TUM. In 2019, she received her Ph.D. degree for her research on proteins in einkorn, emmer, and spelt, with a focus on technological properties and triggers of wheat-related hypersensitivities. The Cereals & Grains Association awarded her work with the Walter Bushuk Graduate Research Award for excellence in cereal protein chemistry. Currently, she works as a postdoctoral researcher in the group of Prof. Katharina Scherf at the Karlsruhe Institute of Technology (KIT). Her research studies the relationship between the structure and functionality of proteins in ancient and modern wheats using modern proteomics methods.
 Katharina A. Scherf leads the Department of Bioactive and Functional Food Chemistry at the Institute of Applied Biosciences, Karlsruhe Institute of Technology (KIT), Karlsruhe, Germany. Together with her team, she studies the complex interplay between structure, functionality, and bioactivity of food biopolymers in a multidisciplinary way and uses these fundamental insights to improve food security, food quality, and food safety. One of her main research interests is analytical, immunological, and biochemical aspects of celiac disease, non-celiac gluten sensitivity, and wheat allergy. Having studied food chemistry, Katharina obtained her Ph.D. degree from the Technical University of Munich and was awarded several scientific prizes, including the Cereals & Grains
Young Scientist Research Award in 2018 and the Gerhard-Billek-Prize from the German Chemical Society in 2015. Katharina A. Scherf leads the Department of Bioactive and Functional Food Chemistry at the Institute of Applied Biosciences, Karlsruhe Institute of Technology (KIT), Karlsruhe, Germany. Together with her team, she studies the complex interplay between structure, functionality, and bioactivity of food biopolymers in a multidisciplinary way and uses these fundamental insights to improve food security, food quality, and food safety. One of her main research interests is analytical, immunological, and biochemical aspects of celiac disease, non-celiac gluten sensitivity, and wheat allergy. Having studied food chemistry, Katharina obtained her Ph.D. degree from the Technical University of Munich and was awarded several scientific prizes, including the Cereals & Grains
Young Scientist Research Award in 2018 and the Gerhard-Billek-Prize from the German Chemical Society in 2015.
References References
- Aprile, M. C., Caputo, V., and Nayga, R. M. Consumers’ preferences and attitudes toward local food products. J. Food Prod. Market 22:19, 2016.
- Avni, R., Nave, M., Barad, O., Baruch, O., Twardziok, S. O., et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 357:93, 2017.
- Blatter, R. H., Jacomet, S., and Schlumbaum, A. About the origin of European spelt (Triticum spelta L.): Allelic differentiation of the HMW glutenin B1-1 and A1-2 subunit genes. Theor. Appl. Genet. 108:360, 2004.
- Borghi, B., Castagna, R., Corbellini, M., Heun, M., and Salamini, F. Breadmaking quality of einkorn wheat (Triticum monococcum ssp. monococcum). Cereal Chem. 73:208, 1996.
- Boukid, F., Folloni, S., Sforza, S., Vittadini, E., and Prandi, B. Current trends in ancient grains-based foodstuffs: Insights into nutritional aspects and technological applications. Compr. Rev. Food Sci. Food Safety 17:123, 2018.
- Brandolini, A., Volante, A., and Heun, M. Geographic differentiation of domesticated einkorn wheat and possible Neolithic migration routes. Heredity 117:135, 2016.
- Catassi, C., Alaedini, A., Bojarski, C., Bonaz, B., Bouma, G., et al. The overlapping area of non-celiac gluten sensitivity (NCGS) and wheat-sensitive irritable bowel syndrome (IBS): An update. Nutrients 9:1268, 2017.
- Fares, C., Codianni, P., Nigro, F., Platani, C., Scazzina, F., and Pellegrini, N. Processing and cooking effects on chemical, nutritional and functional properties of pasta obtained from selected emmer genotypes. J. Sci. Food Agric. 88:2435, 2008.
- Feuillet, C., Langridge, P., and Waugh, R. Cereal breeding takes a walk on the wild side. Trends Genet. 24:24, 2008.
- Fogarasi, A. L., Kun, S., Tankó, G., Stefanovits-Bányai, E., and Hegyesné-Vecseri, B. A comparative assessment of antioxidant properties, total phenolic content of einkorn, wheat, barley and their malts. Food Chem. 167:1, 2015.
- Food and Agriculture Organization of the United Nations, Statistics Division. Food and agriculture data. Published online at http://faostat.fao.org. FAOSTAT, Rome, Italy, 2019.
- Geisslitz, S., Longin, C. F. H., Scherf, K. A., and Koehler, P. Comparative study on gluten protein composition of ancient (einkorn, emmer and spelt) and modern wheat species (durum and common wheat). Foods 8:409, 2019.
- Geisslitz, S., Ludwig, C., Scherf, K. A., and Koehler, P. Targeted LC-MS/MS reveals similar contents of alpha-amylase/trypsin-inhibitors as triggers of non-celiac gluten sensitivity in all wheat species except einkorn. J. Agric. Food Chem. 66:12395, 2018.
- Geisslitz, S., Wieser, H., Scherf, K. A., and Koehler, P. Gluten protein composition and aggregation properties as predictors for bread volume of common wheat, spelt, durum wheat, emmer and einkorn. J. Cereal Sci. 83:204, 2018.
- Giambanelli, E., Ferioli, F., Kocaoglu, B., Jorjadze, M., Alexieva, I., Darbinyan, N., and Antuono, F. A comparative study of bioactive compounds in primitive wheat populations from Italy, Turkey, Georgia, Bulgaria and Armenia. J. Sci. Food Agric. 93:3490, 2013.
- Hidalgo, A., Yilmaz, V. A., and Brandolini, A. Influence of water biscuit processing and kernel puffing on the phenolic acid content and the antioxidant activity of einkorn and bread wheat. J. Food Sci. Technol. 53:541, 2016.
- Koenig, A., Konitzer, K., Wieser, H., and Koehler, P. Classification of spelt cultivars based on differences in storage protein compositions from wheat. Food Chem. 168:176, 2015.
- Longin, C. F. H., and Würschum, T. Back to the future—Tapping into ancient grains for food diversity. Trends Plant Sci. 21:731, 2016.
- Longin, C. F. H., Ziegler, J., Schweiggert, R., Koehler, P., Carle, R., and Wuerschum, T. Comparative study of hulled (einkorn, emmer, and spelt) and naked wheats (durum and bread wheat): Agronomic performance and quality traits. Crop Sci. 56:302, 2015.
- Ludvigsson, J. F., Leffler, D. A., Bai, J. C., Biagi, F., Fasano, A., et al. The Oslo definitions for coeliac disease and related terms. Gut 62:43, 2013.
- Luo, M. C., Yang, Z. L., You, F. M., Kawahara, T., Waines, J. G., and Dvorak, J. The structure of wild and domesticated emmer wheat populations, gene flow between them, and the site of emmer domestication. Theor. Appl. Genet. 114:947, 2007.
- Maccaferri, M., Harris, N. S., Twardziok, S. O., Pasam, R. K., Gundlach, H., et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 51:885, 2019.
- Migliorini, P., Spagnolo, S., Torri, L., Arnoulet, M., Lazzerini, G., and Ceccarelli, S. Agronomic and quality characteristics of old, modern and mixture wheat varieties and landraces for organic bread chain in diverse environments of northern Italy. Eur. J. Agron. 79:131, 2016.
- Molberg, Ø., Uhlen, A. K., Jensen, T., Solheim Flaete, N., Fleckenstein, B., Arentz-Hansen, H., Raki, M., Lundin, K. E. A., and Sollid, L. M. Mapping of gluten T-cell epitopes in the bread wheat ancestors: Implications for celiac disease. Gastroenterology 128:393, 2005.
- Nwaru, B. I., Hickstein, L., Panesar, S. S., Roberts, G., Muraro, A., et al. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy 69:992, 2014.
- Pasqualone, A., Piergiovanni, A. R., Caponio, F., Paradiso, V. M., Summo, C., and Simeone, R. Evaluation of the technological characteristics and bread-making quality of alternative wheat cereals in comparison with common and durum wheat. Food Sci. Technol. Int. 17:135, 2011.
- Rossmann, A., Buchner, P., Savill, G. P., Hawkesford, M. J., Scherf, K. A., and Mühling, K. H. Foliar N application at anthesis alters grain protein composition and enhances baking quality in winter wheat only under a low N fertiliser regimen. Eur. J. Agron. 109:125909, 2019.
- Schalk, K., Lang, C., Wieser, H., Koehler, P., and Scherf, K. A. Quantitation of the immunodominant 33-mer peptide from alpha-gliadin in wheat flours by liquid chromatography tandem mass spectrometry. Sci. Rep. 7:45092, 2017.
- Scherf, K. A. Immunoreactive cereal proteins in wheat allergy, non-celiac gluten/wheat sensitivity (NCGS) and celiac disease. Curr. Opin. Food Sci. 25:35, 2019.
- Shewry, P. R. Do ancient types of wheat have health benefits compared with modern bread wheat? J. Cereal Sci. 79:469, 2018.
- Shewry, P. R., and Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal Sci. 65:236, 2015.
- Shewry, P. R., Piironen, V., Lampi, A.-M., Edelmann, M., Kariluoto, S., et al. The HEALTHGRAIN wheat diversity screen: Effects of genotype and environment on phytochemicals and dietary fiber components. J. Agric. Food Chem. 58:9291, 2010.
- Soriano, J. M., Villegas, D., Aranzana, M. J., Garcia Del Moral, L. F., and Royo, C. Genetic structure of modern durum wheat cultivars and Mediterranean landraces matches with their agronomic performance. PLoS One. DOI: https://doi.org/10.1371/journal.pone.0160983. 2016.
- Souci, S. W., Fachmann, W., and Kraut, H. Food Composition and Nutrition Tables, 8th ed. Wissenschaftliche Verlagsgesellschaft, Stuttgart, Germany, 2016.
- Tatham, A. S., and Shewry, P. R. Allergens to wheat and related cereals. Clin. Exp. Allergy 38:1712, 2008.
- Van den Broeck, H. C., de Jong, H. C., Salentijn, E. M. J., Dekking, L., Bosch, D., Hamer, R. J., Gilissen, L. J. W. J., van der Meer, I. M., and Smulders, M. J. M. Presence of celiac disease epitopes in modern and old hexaploid wheat varieties: Wheat breeding may have contributed to increased prevalence of celiac disease. Theor. Appl. Genet. 121:1527, 2010.
- Vincentini, O., Maialetti, F., Gazza, L., Silano, M., Dessi, M., De Vincenzi, M., and Pogna, N. E. Environmental factors of celiac disease: Cytotoxicity of hulled wheat species Triticum monococcum, T. turgidum ssp. dicoccum and T. aestivum ssp. spelta. J. Gastroenterol. Hepatol. 22:1816, 2007.
- Zentgraf, H. Unser Brotgetreide Nr. 1. (In German) Mehl Rep. 22(2):1, 2013.
- Zevallos, V. F., Raker, V., Tenzer, S., Jimenez-Calvente, C., Ashfaq-Khan, M., Rüssel, N., Pickert, G., Schild, H., Steinbrink, K., and Schuppan, D. Nutritional wheat amylase-trypsin inhibitors promote intestinal inflammation via activation of myeloid cells. Gastroenterology 152:1100, 2017.
|